
Open Access, Volume 9
Evolutionary aspects of a typical pathological process
VP Reutov1*; EG Sorokina2
1Institute of Higher Nervous Activity and Neurophysiology, Russian Academy of Sciences, Russia, 117485, Moscow, st. Butlerova, 5a,
2National Medical Research Center for Children’s Health of the Ministry of Health of Russia, 119991, Moscow, Lomonosovsky Prospekt, 2.
Reutov VP
Institute of Higher Nervous Activity and Neurophysiology, Russian Academy of Sciences, Russia, 117485, Moscow, st. Butlerova.
Email: valentinreutov@mail.ru
Received : March 20, 2023,
Accepted : May 19, 2023
Published : May 23, 2023,
Archived : www.jclinmedcasereports.com
Abstract
Any pathological process proceeds on the background of hypoxia/ischemia, inflammatory, immune or autoimmune processes. These processes lead to an increase in active forms of nitrogen (•NO/•NO2) and oxygen (•O2-/•OH). The article analyzes the relationship between the violation of cyclic regulatory mechanisms and the formation of active forms of nitrogen and oxygen that can interact with each other to form even more reactive compounds that can participate in the oxidation and damage of the main components of cellular structures – proteins, nucleic acids and unsaturated fatty acids. The mechanisms of NO and •O2- cycles that support the production of Reactive Oxygen Species (ROS) and Reactive Nitrogen Species (RNS) within the physiological norm are analyzed. The connection between the ideas of Rudolf Virchow (1821-1902), L.A. Orbeli (1882–1958), G.N. Kryzhanovsky (1922-2013) and the ideas of modern scientists and doctors about principle of cyclicity in space and time can form the foundation for understanding elementary biochemical reactions, the violation of which leads to the development of a typical pathological process (TPP). This TPP according to some scientists and doctors can be considered as a common denominator of all pathological processes occurring on the background of hypoxia / ischemia, inflammatory, activation of immune and autoimmune processes. Excessive production of Reactive Oxygen Species (ROS) and Reactive Nitrogen Species (RNS) can lead to oxidative/nitrosative stress, which is known to damage cells, subcellular structures, organs and tissues. Oxidative and nitrosative stress as a result of the formation of nitrogen dioxide, peroxynitrites, OH-radicals destroys the biochemical structures of the body and leads to a transition from normal physiological processes to pathological processes.
Keywords: Typical pathological process; Nitric oxide cycle; Superoxide anion-radical cycle; Regulation of the content of reactive nitrogen and oxygen species.
Copy right Statement: Content published in the journal follows Creative Commons Attribution License. (http://creativecommons.org/licenses/by/4.0). © Reutov VP (2023)
Journal: Open Journal of Clinical and Medical Case Reports is an international, open access, peer reviewed Journal mainly focused exclusively on the medical and clinical case reports.
Citation: Reutov VP, Sorokina EG. Evolutionary aspects of a typical pathological process. Open J Clin Med Case Rep. 2023; 2042.
Introduction
Nobody knows what life is. No one knows why normal physiological processes turn into pathological processes. Disease, like life itself, cannot be given an exhaustive definition, since disease is a special form of the life process, a qualitatively new form of life activity that occurs in response to an extreme stimulus and manifests itself in distorted regulatory mechanisms and a decrease in adaptability. Identification of regulatory components, analysis and generalization of the mechanisms underlying the transition of normal physiological processes to pathophysiological ones are therefore important tasks of modern biology and medicine. On what factors does human health primarily depend?
Human health, life and its quality are determined by the state of the regulatory systems of cells, organs and the body as a whole. The words of Rudolf Virchow (1821-1902) are still relevant: « It is not life in improper conditions or a distortion alone that causes disease; in contrast, disease starts with a failure of the regulatory system». Analysis of the literature data and the results of our morphological and biochemical studies made it possible to identify the main mechanisms leading to the transition from normal physiological processes to the development of general pathological changes. The review is devoted to the analysis of ideas that arose in the course of solving experimental and clinical problems. Some of these decisions were of great practical importance and significantly affected the life expectancy of the population in the USSR and Russia.
Primary and secondary atmosphere of earth and atoms, that have become the main components of cells
Earth’s primary and secondary atmosphere and the atoms that have become the main components of cells [1,2]. According to modern concepts, most of the Earth’s primary atmosphere consisted of hydrogen (1H2) and helium (4He). Carbon (12C), nitrogen (14N) and oxygen (16O) atoms, in accordance with modern concepts, appeared during the so-called CNO-cycle, proposed and substantiated in 1938 by Bethe in the analysis of three connected and partially overlapping cycles. These events took place in the solar system more 5 billion years ago. At present, such reactions can be observed by modern astrophysical methods in the Andromeda galaxy [3]. In 1967, an American astrophysicist and nuclear physicist Hans Bethe won the Nobel Prize in Physics for «contributions to the verification of nuclear reactions, especially for the discovery of the energy sources of the star».
Thus, according to modern concepts, the primary atmosphere of the Earth was formed from a proto planetary cloud during the formation of the solar system as a result of the capture of the above substances/elements by the gravitational field of the Earth [4,5]. The secondary atmosphere of the Earth, which arose after volcanic eruptions, mainly contained hydrogen (H2), methane (CH4), ammonia (NH3), and interconnected hydrogen and oxygen atoms (H2O) [3–11]. The atoms that were part of the Earth’s primary and secondary atmosphere became the main components of the cells of microorganisms, plants, and animals [10]. Figuratively speaking, nature created life from what was.
It is now known that all living organisms are 96% composed of hydrogen (1H2), carbon (12C), nitrogen (14N) and oxygen (16O) atoms, which were part of the primary and secondary atmosphere of the Earth in a free (H2) state or in a state bound to other atoms (CH4, NH3, H2O ) [11–15]. The rest of the atoms, according to modern concepts, including metals and trace elements that can be found in living organisms, make up no more than 4%. They appeared, as some researchers suggest, from the bowels of the planet as a result of volcanic eruptions [1-8]. The question of how inorganic compounds and organic substances entered the composition of living organisms, realizing numerous regulatory mechanisms and systems at various structural and functional levels, is the subject of research for the entire set of biomedical disciplines [9-12]. Why this became possible, and why nature is the same at all structural and functional levels, remains one of the mysteries that physicists, chemists and biologists have been trying to solve over the past centuries [8,14,16]. However, these questions, formulated in a different form, have worried almost all natural scientists in the world, at least for 2500 years. In this article, analyzing evolutionary aspects of a typical pathological process together with the evolution of the energy apparatus and problem of gas transmitters in mammals, we will try to give one of the possible answers to the questions posed.
The evolution of the energy apparatus of cells
According to existing ideas, the driving force behind phylogenetic metamorphosis, at least in the early stages of development, was the evolution of energy metabolism, regulation and storage of energy [17-19]. Energy metabolism in the earliest forms of life appears to have been neither photosynthetic nor aerobic. It could be similar to the glycolytic pathway, which is known to be carried out during substrate phosphorylation, but without the electron transport characteristic of mitochondria [17-21]. However, it should be noted that glycolysis, despite its relative “simplicity”, requires the participation of enzymes, therefore, it needs the mechanisms of protein synthesis [20-22]. In this regard, there is every reason to believe that, despite the relative simplicity of glycolysis, this energy mechanism cannot be considered primitive. The modern system of protein biosynthesis, which is under genetic control, is a very complex mechanism. This mechanism does not allow the reconstruction of amino acid and nucleotide polymerization systems.
The History of Oxygen on Earth is the History of Life, Disease and Death of Living Organisms. Oxygenic phototrophs have played a fundamental role in Earth’s history by enabling the rise of atmospheric oxygen (O2) and paving the way for animal evolution [23]. The formation of organisms, as mentioned above, on the planet Earth took place 4–5 billion years ago. At the same time, there was no oxygen (O2) on our planet for about 2 billion years. Instead of oxygen, nitrates/nitrites and sulfates/sulfites were used as electron acceptors. In addition, the microorganisms living in the mines developed a type of respiration, which was later called carbamate. This type of respiration is still used by methane-producing microorganisms [26,27]. Why were these low molecular weight compounds (NO3-/NO2-; SO4-/SO3) - necessary for living organisms?
First of all, these substances or compounds, which were used by living organisms leading a sedentary lifestyle, were necessary to meet energy needs. Thus, for example, nitrite ions were used by the electron transport chains (or respiratory chains) of mitochondria as terminal (or terminal) electron acceptors [28-38]. Then, when oxygen appeared, the same electron transport chains were adapted for the transfer of electrons to O2 [28]. The realization of this fact led scientists to put forward a hypothesis that nitrate-nitrite respiration was a precursor of oxygen respiration, and mitochondria in eukaryotic cells appeared as endosymbionts (endosymbiotic hypothesis of the origin of mitochondria) [17,18]. Plants and microorganisms that widely use nitrates and nitrites as electron acceptors had a low-energy metabolism [31-33]. Animals, fish and birds that had significantly higher metabolisms were able to run, swim or fly. However, with this metabolism, they produced life-threatening free oxygen radicals, which, after interaction with free nitrogen radicals, led to the formation of nitrogen dioxide (•NO2), peroxynitrites, •OH radicals [39,40]. This led to a reduction in life expectancy: life became more interesting, but much shorter. In addition, diseases that plants and microorganisms did not know were added at a relatively early age. Everything has to be paid for, including life and health. No wonder they say that the history of oxygen on Earth is the history of life, disease and death of living organisms.
Cyclic organization of reactive nitrogen specias (RNS) and reactive oxygen specias (ROS)
Compounds were formed with the participation of hydrogen, carbon, nitrogen, and oxygen atoms, which began to perform an energetic function under the conditions of the existence of unicellular organisms [9-12]. The same atoms laid the foundation for Nitric Oxide (NO), superoxide radical anion (•O2-), carbon monoxide (CO), as well as hydrogen sulfide (H2S), sulfur dioxide (SO2) and polysulfides (H2Sn), which began to function as intracellular mediators [13-40]. After the emergence of multicellular organisms, compounds SO3 to SO3-: •O2-/O2, NO3-/NO2-, SO4-/SO3-, CO2/CO, which were previously formed during respiration, were used as electron acceptors, or in intracellular signaling systems, were adapted for intercellular signaling [41-48].
However, these processes and phenomena were unknown in science in the second half of the XX century. Before the discovery of these compounds in living organisms, the discovery of their endogenous production, these molecules were known as toxic substances. Therefore, we can say with good reason that the problem of gaseous intermediaries (gas transmitters) has become a new problem in the 21st century [42-52]. It is in recent decades that scientists have come to realize the specific role of gas transmitters that perform a signaling function in cells [15-20]. These substances have been assigned to a new class of biologically active substances [37-52]. Gas transmitters are small molecules that easily cross biological membranes and do not act through special receptors [43-53]. These compounds are synthesized in the body using special enzymes and are labile (their half-life is measured in seconds). They do not accumulate in cells and subcellular structures, since they cannot be enclosed in vesicles, like neurotransmitters [43-51]. Sometimes they can be bound by metals of proteins and enzymes, as well as SH-groups that are part of amino acids, bio phenols, for example, tyrosine and its derivatives, free or part of proteins, etc. Gas transmitters have endocrine, paracrine, autocrine, and autocrine, and intracrine effects [42-52].
Cyclic organization of gas transmitters: Cycles of Nitric Oxide and Super Oxide Anion-Radical
Back in the 1970s one of the authors of this work (V.P.R) drew attention to cyclic processes involving active forms of nitrogen and oxygen. Cyclic changes in the concentration of NO and •O2 in various tissue cells and in the whole organism suggested that these active compounds are controlled by NO and •O2- cycles and are an essential part of the cell regulatory mechanisms of tissues of living organisms that protect cells and the body as a whole from the development of oxidative and nitrosative stress. An analysis of the literature and the results of our own studies led to the formulation and substantiation of the concepts of the nitric oxide cycle and the superoxide anion-radical [54-56]. The results of these studies and their generalizations were reported in Stockholm at the Karolinska Institute and included in a paper published in Nature Chemical Biology [58].
A small increase in physiological nitrite concentrations has been found to mediate a number of biological responses, including hypoxic vasodilation, cytoprotection after ischemia/reperfusion, and regulation of gene and protein expression. Thus, while nitrite was considered biologically inert until recently, it is now recognized as a potentially important hypoxic signaling molecule and therapeutic agent. Nitrite mediates signaling through its reduction to nitric oxide through reactions with several heme-containing proteins. It has been established that cytochrome c and cytochrome oxidase (cyt a + a3) can play an important role as a nitrite reductase that converts nitrites into NO [29,59,60]. An important role in substantiating the concept of nitric oxide and superoxide anion radical cycles was played by numerous open nitrite reductase reactions involving the heme-containing proteins of hemoglobin, myoglobin, cytochrome oxidase (cyt a + a3), and cytochrome P-450, which, under conditions of hypoxia/ischemia, are capable of converting ions NO2- to NO [29,59-63]. Together with NO synthase reactions, they completed the chain of nitrite reductase reactions and led to the substantiation of the concept of the nitric oxide cycle and the superoxide anion radical [54-58].
An analysis of the mechanisms of nitric oxide and superoxide anion radical cycles made it possible to establish a connection between the ideas of R. Virkhow, L.A. Orbeli, G.N. Kryzhanovsky and modern scientific doctors [64-68]. L.A. Orbeli said: “We take little into account the fact that all physiological processes are carried out cyclically and each process has its own cyclicity.” He often said: “We often ignore the fact that all processes are cyclic and each process has its own cycle.” His words echoed the ideas of R. Virchow” It is not living in unsuitable conditions that causes disease; on the contrary, the disease begins with a failure of the regulatory system. It becomes clear the relationship between the ideas of L.A. Orbeli and R. Virchow [64-68]. To do this, it is necessary to take into account the fact that all regulatory systems operate in a cyclic mode. G.N. Kryzhanovsky came to independent conclusions: every pathology is «a dysregulatory pathology». The mechanisms of NO and O2- cycles supports the production of reactive oxygen species (ROS) and Reactive Nitrogen Species (RNS) within the physiological norm. The analysis made it possible to isolate the main mechanisms that underlie a transition from normal physiological processes to common pathological changes.
Scheme of nitric oxide (NO) and superoxide anion radical (•O2-) cycles
The concept of the nitric oxide cycle (Figure 1a) was substantiated 29 years ago [69]. Its substantiation became possible due to the detection of nitrite reductase activity of heme-containing proteins in mammals [70]. After 20 years, the nitrite reductase activity of hemoglobin in the deoxy-form was confirmed in the works of researchers from the United States [71]. Subsequently, this concept has repeatedly found its development and application for various normal and pathological processes [28,37-52,72].
However, in the simplest form, we substantiated this concept in the mid-70s of the 20th century. Thanks to the support of the Academy of Sciences of the USSR and the Russian Academy of Sciences, it was possible to carry out a number of biomedical measures in the country, which increased the average life expectancy of the population of the USSR by 5-6 years. This achievement was included in the Reports of the Department of Physiology of the USSR Academy of Sciences, and the authors received awards – one gold medal, one silver medal and three bronze medals (1983). It is described in more detail in the section «Results of Practical Application of Concept of NO and •O2- Cycles and Typical Pathological Process».
The essence of this concept is that NO-2- ions formed from L-arginine can again, with the participation of nitrite reductase systems, including Hb, Mb, cyt a + a3 and cyt P-450, close the chain: L-arginine → NO → NO2-/NO3- into a cycle. Oxygen, binding with heme, inhibits the nitrite reductase activity of these proteins [70,71]. Thus, at various functional states associated with insufficient oxygen supply to the body, the nitrite reductase component of the nitric oxide cycle will be activated (Figure 1a) [28,54-56].
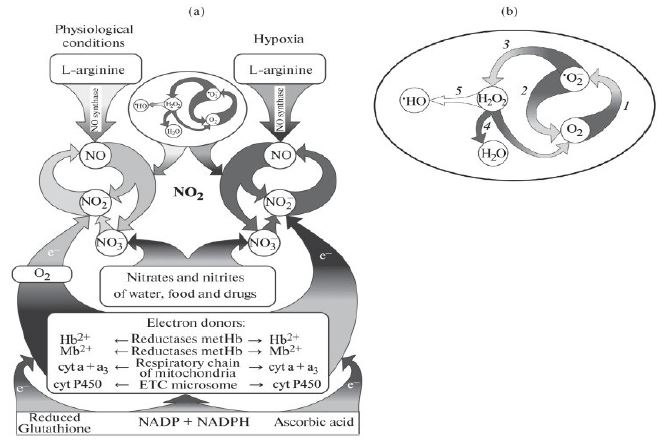
Figure 1: Cycle of nitric oxide (a) and superoxide radical anion (b).
In the nitric oxide cycle, one can distinguish the NO-synthase component (“L-arginine - NO”), which synthesizes NO in the presence of oxygen, and the nitrite reductase component, the activity of which sharply increases under conditions of oxygen deficiency (hypoxia/ischemia). The formation of NO with the participation of the NO-synthase component is carried out as a result of the oxidation of the guanidine nitrogen of L-arginine. NO2-, ions, formed from L-arginine, can again, with the participation of nitrite reductase systems, including Hb, Mb, cyt a + a3 and cyt P-450, close the chain of L-arginine → NO → NO2-/ NO3- into a cycle. Oxygen, binding to heme, inhibits the nitrite reductase activity of these proteins. During hypoxia and functional stress, when heme-containing proteins are converted into deoxy-form, NO2- ions begin to actively reduce, accepting electrons from these heme-containing proteins. An important role in the reduction of NO2– ions to NO is also played by electron donor systems that reduce Hb, Mb, cyt a + a3, and cyt P-450. In addition to these electron transport systems (chains), ascorbic acid and reduced glutathione can play an important role in the reduction of NO2- ions. In the cycle of superoxide radical anion occur: 1 – reduction of oxygen (O2) and the formation of superoxide anion radical (•O2-); 2 and 3 – superoxide dismutation reactions catalyzed by superoxide dismutase; 4 – decomposition of hydrogen peroxide (H2O2) into water (H2O) and molecular oxygen (O2), carried out by the enzyme catalase; 5 – hydrogen peroxide (H2O2) 5 – hydrogen peroxide – H2O2 – also decomposes to form two molecules of the •OH-radical. The cyclic organization of reactive nitrogen (•NO2) and oxygen (•O2-) species ensures the conversion of these reactive, highly reactive compounds into less active substances. When the cycles of nitric oxide and superoxide anion-radical are disrupted, even more active molecules of nitrogen dioxide (•NO2) and peroxynitrites appear, again decaying into •NO2 and •OH-radicals, which damage the main components of living organisms.
However, such activation can be observed until the depletion of L-arginine occurs [66]. L-arginine is included in the Krebs cycle at the level α-ketoglutarate complex and serves as one of the sources of formation of succinate (or succinic acid). During relatively prolonged hypoxia / ischemia, as is known, succinate is actively used as an oxidation substrate for the formation of ATP and membrane potential in mitochondria.
Analysis of the literature data and the results of our own research allowed us to put forward a hypothesis that, in addition to the nitric oxide cycle, there should also be a cycle of superoxide anion radical (Figure 1b) [55]. Oscillations in the concentrations of reactive oxygen species in biological systems, obtained by a number of authors, testified in favor of the existence of such a cycle. Since all cyclic processes always involve periodic oscillations, one could expect that the previously considered reaction products associated with the neutral O2 molecule and its active forms - superoxide, peroxide, as well as enzymes for activating molecular oxygen (Fe2+- and Сu2+- containing proteins), superoxide dismutase and catalase can be closed in a cycle. The analysis of numerous literature data allowed us to propose a scheme for the cyclic organization of reactive oxygen species, which we called the superoxide anion radical cycle (Figure 1b) [74].
Scheme of hydrogen sulfur / sulfur dioxide cycle (H2S / SO2)
We summarized numerous literature data on gas transmitters (NO, •O2-, H2S/H2Sn/SO2) and formulated the concept of cyclic transformations Hydrogen Sulfur / Sulfur Dioxide Cycle (Figure 2). The initial substrate for the synthesis of all three mediators (H2S/H2Sn/SO2) is sulfur-containing amino acids, primarily L-cysteine. Polysulfides are formed from hydrogen sulfide (H2S), and sulfur dioxide (SO2) is sulfite anhydride (sulfurous acid), which is formed during the oxidation of Н2S [47]. The end product of the oxidation (degradation) of Н2S, like the oxidation of sulfur dioxide, is sulfate, which is excreted in the urine. It is known that the intestinal microflora synthesizes hydrogen sulfide from sulfate, reducing it. It is characteristic that an enzyme that oxidizes Н2S in mitochondria in animals, sulfide quinone oxidoreductase, SQR, was also found in plants and bacteria [47]. The reduction of sulfate (SO4) to Н2S is possible in mammals with the participation of intestinal microflora. Thus, the cycle of hydrogen sulfide in the whole body of mammals exists (Figure 2). Our assumption about the reduction of SO4 to Н2S in mammalian cells is hypothetical. If such a reaction occurs, then we can say that the hydrogen sulfide cycle exists not only in the whole organism of mammals, but also in their cells. The cycles presented above are, from our point of view, a manifestation of the global principle of cyclicity [54-56,72]. At the same time, the geochemical cycles of nitrogen, carbon, sulfur and water are known as the natural cycles. Together with the cycles we are analyzing, they are a manifestation of the principle of holography, when the cyclic properties of the general are manifested in the cyclic properties and regularities of the particular, which lies at other structural and functional levels of animate and inanimate nature.
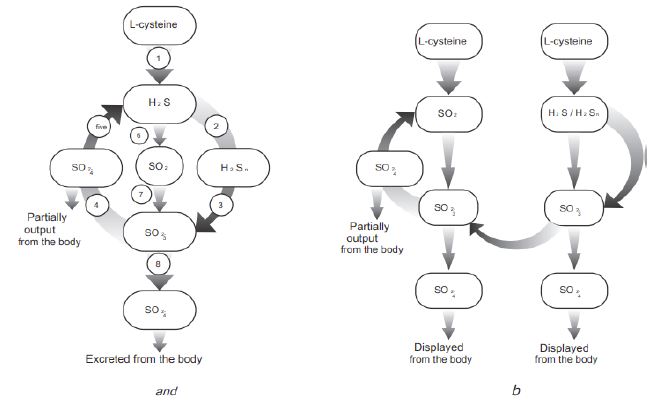
Figure 2: Cycle of hydrogen sulfide (a) and its SO2- and H2S/H2Sn compounds (b), which can be activated at various functional states.
The initial substrate for the synthesis of all S-mediators is sulfur-containing amino acids, primarily L-cysteine. Hydrogen sulfide (H2S) is synthesized from L-cysteine (stage 1). Currently, 3 enzymes are known that are involved in the synthesis of H2S at this stage: in various mammalian cells: cystathionine- β-synthase (CBS), Cystathionine-γ-Lyase (CSE) and 3-mercaptopyruvate sulfurtransferase (3-MST). CBS synthesizes H2S primarily in nerve cells. CSE synthesizes H2S in blood vessel smooth muscle cells, and 3-MST synthesizes H2S in endothelial cells. Polysulfides (H2Sn) are formed from hydrogen sulfide (H2S) (stage 2), and sulfur dioxide (SO2) is formed by the oxidation of H2S or L-cysteine (stage 6). The synthesis of SO2 from L-cysteine occurs under the influence of the enzymes cysteine dioxygenase and aspartate aminotransferase (stage 7). Further, as H2S, so SO2 is converted to sulfate (SO42- ). An intermediate stage of this transformation for both H2S and SO2 is sulfite (SO32-). However, H2S is initially oxidized to persulfide (RSSH) or polysulfide (H2Sn) (stage 2) under the influence of sulfide quinone oxidoreductase (SQR), which is oxidized by persulfide oxygenase (ETEH1) to sulfite (stage 3). Sulfur dioxide (SO2) can be directly hydrogenated to sulfuric acid (H2SO3) to form sulfite ions (SO32-) (step 8). The latter is oxidized by sulfite oxidase to sulfate (SO42-) (stage 4), which, entering the intestine, is reduced to H2S (stage 5) and excreted in the urine. The end product of H2S oxidation, as well as sulfur dioxide (SO2) oxidation, is sulfate (SO42-), which either enters the intestines and is partially excreted from the body, or is excreted with urine. It is known that the intestinal microflora reduces SO42- with the formation of H2S. It is characteristic that the SQR enzyme, which oxidizes H2S in animal mitochondria, is also found in plants and bacteria. The reduction of sulfate (SO4) to H2S is possible in mammals with the participation of intestinal microflora. Thus, the cycle of hydrogen sulfide in the whole body of mammals exists. Our hypothesis on the reduction of SO42- to H2S in mammalian cells is hypothetical. If such a reaction occurs, then we can say that the H2S cycle exists not only in the whole organism of mammals, but also in their cells.
Typical Pathological Process
Any pathological process proceeds on the background of hypoxia / ischemia, inflammatory, immune or autoimmune processes. These processes lead to an increase in Reactive Nitrogen Species (RNS) and Reactive Oxygen Species (ROS). Our generalizing concept considers of how the pathological process develops. Following the concept, the typical pathological process is based on nonspecific distortion of cyclic regulatory processes and arises when RNS and ROS increase simultaneously [74]. Once RNS and ROS concentrations are beyond the regulatory capabilities of biochemical antioxidant systems, nitric oxide and superoxide anion radical cycles are disrupted. In the context of the concept, damage to cell membranes and subcellular structures arises because the above alterations lead to the generation of nitric dioxide (NO2), which is a highly reactive compound, is involved in free radical chain reactions, and oxidizes the main biochemical components of living organisms: DNA/RNA (guanines primarily), fatty acids (unsaturated fatty acids that are components of phospholipid membranes), and proteins (the SH groups of sulfur-containing amino acids and the OH groups of tyrosine residues to produce nitrotyrosine). Our generalizing concept agrees well with the ideas that every disease starts with a failure of regulatory mechanisms (R. Virkhov) and that dysregulatory pathology forms its basis (G.N. Kryzhanovsky).
Our concept of a typical pathological process as a common denominator of all pathological processes occurring against the background of hypoxia/ischemia, activation of inflammatory responses, immune and autoimmune processes, and diseases can be extended to many known diseases [74]. Their number may be about 10,000 known pathological processes. The fundamental nature of this concept lies in the fact that it combined the deep processes associated with respiration and bioenergetics, which are characteristic of all living organisms without exception.
Causes and effects of universal principles
Over the past decades, we have suggested that the cyclic organization NO and •O2- in cells and in the whole organism may be a consequence of the existence of such a principle, the universality of which is comparable to the principle of the atomic structure of matter [54-56,72,74]. This principle extends its influence to almost all structural and functional levels in animate and inanimate nature, subordinates the behavior of living subjects and inanimate objects, and, sets the rules for the functioning of all regulatory systems that contain elements of negative and positive feedback. Moreover, this principle explains the nature of positive and negative feedbacks in living organisms and nonliving systems. It also answers the question of why development is carried out in a spiral, and why spirals are one of the main structural elements in DNA, proteins, in nerve fibers (myelin sheaths), glial cells (glial wraps in the central nervous system), etc. This principle also explains the nature of the cyclical / periodic pattern in the functioning of regulatory systems at almost all structural and functional levels in living organisms [72]. In addition, this principle and the consequences arising from it are in good agreement with other concepts and theories of scientists.
Thus, studying very specific substances (nitrates / nitrites and nitrogen oxides), we came to the cyclic organization of their functioning [56–70], then found the same cyclic organization in the systems of other gas transmitters, analyzed the functioning of these compounds in health and development pathological processes [22,72]. These concepts developed both for gas transmitters and for other processes and phenomena of animate and inanimate nature, are fundamentally new. At the same time, they are in good agreement with practically all other known experimental data and theoretical constructions. Let’s give one more example. French mathematician, physicist and philosopher Henri Poincare (1854-1912) created a new branch of mathematics – the qualitative theory of differential equations (1881-1882). He showed the existence of a huge class of phenomena that obey periodic laws. He also answered the question of how, without solving equations, one can obtain important information about the behavior of a family of solutions. “Any generalization to a certain extent presupposes belief in the unity and simplicity of nature. As far as unity is concerned, scientists usually do not encounter any difficulties. The question is, how is nature one? “How is nature united?” (H. Poincare).
From our point of view, the atomic principle of the structure of matter, as well as the principle of cyclicity and the holographic principle proposed and substantiated as a global principle, answer the question of A. Poincare: how is nature united? Let us recall that the holographic principle got its name from the Greek word holos – all, complete and grapho - I write, I draw. The result is: the optical equivalent of the object, the ideas of which were formulated by the Hungarian physicist Denis (Denesh) Gabor (1900–1979) in 1948 with the improvement of the electron microscope [56–60]. H. Poincare realized that behind the numerous periodic processes that he described and analyzed using a system of differential equations, there are global laws. However, he could not know about the existence of the principles of cyclicity and the principle of holography.
There, after posing the question and formulating the problem, A. Poincare left its solution to future generations of researchers. The principle of cyclicity in combination with the holographic principle means that cyclic properties (or properties of a spiral in space, and periodicity in time) should be repeated at all structural and functional levels. In the same way, like the atomic principle of the structure of matter, it should also manifest itself at all structural and functional levels, because these principles are universal or global [75-80]. Thus, the above three principles: the atomic principle of the structure of matter, the principle of cyclicity and the holographic principle, structure and combine the phenomena of animate and inanimate nature. It is these three principles, from our point of view, that make nature one (or unit), including inorganic compounds and organic substances into a single system. The cyclic organization of gas transmitters in mammals is one of the manifestations of this unity [81]. At present the ideas of cyclicity, periodicity, and holography are increasingly being used in physiology, pathophysiology, and medicine [75-80].
To say that this is philosophy and not biochemistry, biophysics, physiology or medicine, from our point of view, is inappropriate. Good, deep and concrete science always has a direct connection with the philosophy of natural science. In the same way, like the philosophy of natural science, finds its embodiment and manifestation in specific sciences. Scientists from different countries have always strived to achieve the unity of the philosophy of natural science and specific sciences. Deep concrete science always has a direct connection with the philosophy of natural science.
Results of practical application of concept of NO and •O2- cycles and typical pathological process
These ideas about the protective role of nitric oxide cycles and the cycle of superoxide anion radical (in the simplest form) were supported for their further implementation in solving practical problems of health care. The fact is that there was a decrease in the average life expectancy for 18 years in the USSR (1962-1980) (Figure 3).
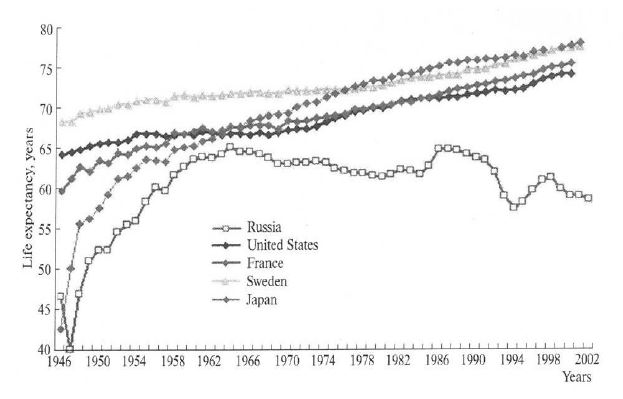
Figure 3: Life expectancy in the USSR/Russia, USA, France, Sweden and Japan (1946–2002).
Source: Human Mortality Database, http://c.avsim.su/i?u=http://www.demoscope.ru/weekly/2004/0169/img/t_graf01_1.gif.
The reasons for this phenomenon were unknown. Therefore, at the first stage, they tried to overcome the decline in average life expectancy in the USSR by increasing the number of doctors, hospitals and polyclinics (Table 1). For 69 years in the USSR (1922-1991), the number of doctors in the country increased by more than 60 times, from 10.9 thousand in 1921 to 667.3 thousand in 1990. Before the War (1939/1941-1945), there were 7.4 doctors per 10000 people, before the collapse of the USSR – 45. In the 1970s, after the number of doctors in the USSR (1/6 of the land) began to exceed all conceivable limits (about 20–25% of world indicators) (Table 1) [82].
Table 1: The number of doctors in the USSR and other countries (generalizing analysis according to data presented on Internet sites) [82].
| Years | Number of doctors of all specialties | ||
|---|---|---|---|
| total, thousand people | per 10 000 people | ||
| USSR | 1966 | 577.7 | 24.6 |
| Russian SSR | 328.3 | 25.8 | |
| Ukrainian SSR | 114 | 24.8 | |
| Byelorussian SSR | 19.8 | 22.6 | |
| Uzbek SSR | 19.5 | 17.9 | |
| Kazakh SSR | 23.4 | 18.9 | |
| Georgian SSR | 16.4 | 35.5. | |
| Azerbaijan SSR | 11.6 | 24.1 | |
| Lithuanian SSR | 7 | 23.1 | |
| Moldavian SSR | 6.3 | 18.5 | |
| Latvian SSR | 7.5 | 32.6 | |
| Kirghiz SSR | 5.3 | 19.4 | |
| Tajik SSR | 4.1 | 15.4 | |
| Armenian SSR | 6.3 | 28.1 | |
| Turkmen SSR | 4.2 | 21.4 | |
| Estonian SSR | 4 | 30.7 | |
| England | 1963 | -79.1 | -14.7 |
| India | 1961 | 83.3 | 1.9 |
| Iran | 1964 | 8.4 | 3.7 |
| Italy | 1961 | 81.2 | 16.3 |
| Pakistan | 1960 | 8.7 | 0.9 |
| United States | 1964 | 360.3 | 18.6 |
| Turkey | 1963 | 10.1 | 3.3 |
| Federal Republic of Germany | 1965 | 110.4 | 19.3 |
| France | 1964 | 75.2 | 15.4 |
| Japan | 1964 | 139.6 | 14.3 |
The problem of reducing life expectancy in the USSR continued to worsen, fundamentally new ideas were required, the implementation of which was noted as one of the most striking achievements of the Institute of Higher Nervous Activity and Neurophysiology of the USSR Academy of Sciences/RAS. The essence of this achievement was that the decrease in the nitrate-nitrite background, the nitrite reductase activity of the enzymatic systems of mammals, and with them the concentrations of NO and NO2 in individuals of the population in the USSR, led to a halt in the catastrophic decline in the average life expectancy of the population of the USSR (Figure 3) [88]. Subsequently, this marked the beginning of the growth of this indicator in the period 1980–1990 for 3–5 years in different regions of the country. After solving this problem, it became clear that the decision of leader of the country USSR and General Secretary of the CPSU – Nikita S. Khrushchev (1894-1971) in May 1957 to increase the yield of agricultural crops – a fodder base for small animals, cattle and poultry-in order to catch up and overtake the United States in the production of meat, butter and milk through chemicalization of agriculture (nitrogen fertilizers), became the main reason for the decrease in the average life expectancy in the USSR. This, in the form of increased chemical load of nitrates and nitrites in water and food, as well as nitrogen oxides in the air, affected health, life expectancy and demographics for at least 18 years (with a latency period of 5 years).
Starting from 1985, this problem began to be actively solved at the Research Institute of Carcinogenesis of the USSR Academy of Medical Sciences/RAMS, Oncological Research Center of the USSR Academy of Medical Sciences/RAMS (now N.N. Blokhin National Medical Research Center of Oncology of the Ministry of Health of Russia), and 6 years later (1989–1992) – at the Institute of Nutrition of the USSR Academy of Medical Sciences/Russian Academy of Sciences and other institutes of the Russian Academy of Sciences and Russian Academy of Medical Sciences [95].
Ultimately, the ideas expressed and the mechanisms discovered by one of the co-authors (V.P.R.) of the article contributed to the fact that in the difficult years of Perestroika, low oil prices, and empty shelves in the shops of the USSR, the population increased by 24.1 million people over 10 years (1980–1990) (from 264.5 million people in 1980 to 288.6 million people in 1990). By the beginning of 1991, the population (according to the current estimate) had increased to 289.2 million people– it was the highest population growth over the past 100 years (1922-2022), and the implementation of this project turned out to be the cheapest project in all the years of the existence of the Academy of Sciences of the USSR/RAS (payment of a scholarship to one graduate student – V.P. Reutov).
Conclusion
The questions of what life is, what is its essence, and what mechanisms and principles underlie living activity of organisms still lack exhaustive answers and definitions that would satisfy all scientists. The only approach today is to list the features that distinguish living systems from inanimate matter. The features are numerous and vary from reproduction to learning. Which one is most important among them? There is still no decisive answer to the question.
Any pathological process proceeds on the background of hypoxia / ischemia, inflammatory, immune or autoimmune processes. These processes lead to an increase in active forms of nitrogen (•NO/•NO2) and oxygen (O2-/•OH). The article analyzes the relationship between the violation of cyclic regulatory mechanisms and the formation of active forms of nitrogen and oxygen that can interact with each other to form even more reactive compounds that can participate in the oxidation and damage of the main components of cellular structures – proteins, nucleic acids and unsaturated fatty acids.
For several centuries, scientists and doctors have tried to identify specific changes in cells, subcellular structures and membranes in each pathological process and disease. However, it turned out that in many pathological processes, first of all, universal nonspecific changes in cells, plasma membranes and membranes of subcellular structures are observed. This made it possible to speak about the existence of typical nonspecific disorders of various cellular and subcellular structures. The concept we are developing about the cyclical organization of gas transmitters (NO, O2-, H2S/SO2) in the norm and its violation during the development of pathological processes makes it possible to understand the mechanisms of development of a typical pathological process, which is the common denominator of almost all pathological processes.
For at least half a century, almost all scientists, including physicists, chemists and biologists working in the field of free radical processes in biology and medicine, have been trying to solve the riddle: how living systems provide regulation and stabilization of reactive oxygen, nitrogen, and also other free radicals. Our proposed concept for the cyclic organization of gas transmitters (NO, O2- and H2S/SO2) may be one of the answers to the question posed more than half a century ago. According to the developed concepts, the above principles make it possible to answer the question: how nature is unified, and how living organisms ensure the regulation and stabilization of nitric oxide (NO), superoxide anion-radical (•O2-) and hydrogen sulfide/sulfur dioxide (H2S/SO2), involved in intra- and intercellular signaling in mammals.
It can be expected that the awareness of the connection between the ideas of R. Virchow (1821-1902), L.A. Orbeli (1882-1958), G.N. Kryzhanovsky (1922-2013) and our ideas about the trinity of the principles of cyclicity, the holographic principle and the principle of the atomic structure of matter allow to answer the question of A. Poincaré: how does nature maintain unity? These ideas and ideas about NO and •O2- cycles form the basis for understanding elementary biochemical reactions, the violation of which leads to the development of a typical pathological process, which, from our point of view, can be considered as a common denominator of all pathological processes occurring on the background of hypoxia / ischemia, inflammation, activation of immune and autoimmune processes.
Acknowledgement: This work was supported by the Russian Academy of Sciences
Compliance with ethical standards
The authors declare that they have no conflict of interest.
This article does not contain any studies, involving animals or human participants performed by any of the authors.
References
- Oka T. H3+, the ideal probe for in situ measurement of the Galactic cosmic rays. Philos Trans A Math Phys Eng Sci. 2019; 377: 20180402.
- Ceccarelli C, Favre C, Lopez-Sepulcre A, Fontani F. Hydrogen molecular ions and the violent birth of the Solar System. Philos. Trans A Math Phys Eng Sci. 2019; 377: 20180403.
- Darnley MJ, Hounsell R, O’Brien TJ, Henze M, Rodriguez-Gil P. A Recurrent Nova Super-Remnant in the Andromeda Galaxy. Nature. 2019; 565: 460-463.
- Kasting JF. Early earth: faint young sun redux. Nature. 2010; 464: 687-689.
- Francl M A brief history of water. Nat Chem. 2016; 8: 897-898.
- Taverne YJ, Merkus D, Bogers AJ, Halliwell B, Duncker DJ. Reactive Oxygen Species: Radical Factors in the Evolution of Animal Life: A molecular timescale from Earth’s earliest history to the rise of complex life. BioEssays. 2018; 40.
- Lyons TW, Reinhard CT, Planavsky NJ The rise of oxygen in Earth’s early ocean and atmosphere. Nature. 2014; 506: 307-315.
- Sánchez-Baracaldo P, Ridgwell A, Raven JA. A neoproterozoic transition in the marine nitrogen cycle. Curr Biol. 2014; 24: 652-657.
- Braakman R. Evolution of cellular metabolism and the rise of a globally productive biosphere. Free Radic Biol Med. 2019; 140: 172-187.
- Stanley SM. Estimates of the magnitudes of major marine mass extinctions in earth history. Proc Natl Acad Sci. 2016; 113: E6325-E6334.
- Braakman R. Evolution of cellular metabolism and the rise of a globally productive biosphere. Free Radic Biol Med. 2019; 140:172-187.
- Kasting JF, Siefert JL. Life and the evolution of Earth’s atmosphere. Science. 2002; 296: 1066-1068.
- Des Marais DJ. Earth’s early biosphere. Gravit Space Biol Bull. 1998; 11: 23-30.
- Nisbet EG, Sleep NH. The habitat and nature of early life. Nature. 2001; 409: 1083-1091.
- Daniel I, Oger P, Winter R. Origins of life and biochemistry under high-pressure conditions. Chem Soc Rev. 2006; 35: 858-875.
- Chu M, Zhang X. Alien species invasion of deep-sea bacteria into mouse gut microbiota. J Adv Res. 2023; 45: 101-115.
- Broda E. The evolution of bioenergetic processes. Prog Biophys Mol Biol. 1970; 21: 143-208.
- Margulis L. Symbiosis and evolution. Sci Am. 1971; 225: 48-57.
- Stojanović O, Miguel-Aliaga I, Trajkovski M. Intestinal plasticity and metabolism as regulators of organismal energy homeostasis. Nat Metab. 2022; 4: 1444-1458.
- Vaupel P, Multhoff G. Revisiting the Warburg effect: historical dogma versus current understanding. J Physiol. 2021; 599: 1745-1757.
- Makinde E, Ma L, Mellick GD, Feng Y. Mitochondrial Modulators: The Defender. Biomolecules. 2023; 13: 226.
- Wu YT, Wu SB, Wei YH. Metabolic reprogramming of human cells in response to oxidative stress: implications in the pathophysiology and therapy of mitochondrial diseases. Curr Pharm Des. 2014; 20: 5510-5526.
- Sánchez-Baracaldo P, Cardona T. On the origin of oxygenic photosynthesis and Cyanobacteria. New Phytol. 2020; 225: 1440-1446.
- Villemur R, Lanthier M, Beaudet R, Lépine F. The Desulfitobacterium genus. FEMS Microbiol Rev. 2006; 30: 706-733.
- Ueno A, Tamazawa S, Tamamura S, Murakami T, Kiyama T, et al. Desulfovibrio subterraneus sp. nov., a mesophilic sulfate-reducing deltaproteobacterium isolated from a deep siliceous mudstone formation. Int J Syst Evol Microbiol. 2021; 71: 4683.
- Pozuelo M, Panda S, Santiago A, Mendez S, Accarino A, et al. Reduction of butyrate- and methane-producing microorganisms in patients with Irritable Bowel Syndrome. Sci Rep. 2015; 5:12693.
- Yang X, Liang Q, Chen Y, Wang B. Alteration of Methanogenic Archaeon by Ethanol Contribute to the Enhancement of Biogenic Methane Production of Lignite. Front Microbiol. 2019; 10: 2323.
- Reutov VP, Sorokina EG. NO-synthase and nitrite-reductase components of nitric oxide cycle. Biochemistry (Mosc). 1998; 63: 874-84.
- Kozlov AV, Staniek K, Nohl H. Nitrite reductase activity is a novel function of mammalian mitochondria. FEBS Lett. 1999; 454: 127-130
- Nohl H, Staniek K, Sobhian B, Bahrami S, Redl H, et al. Mitochondria recycle nitrite back to the bioregulator nitric monoxide. Acta Biochim Pol. 2000; 47: 913-921.
- Takaya N, Kuwazaki S, Adachi Y, Suzuki S, Kikuchi T, et al. Hybrid respiration in the denitrifying mitochondria of Fusarium oxysporum. J Biochem. 2003; 133: 461-465.
- Shoun H, Tanimoto T. Denitrification by the fungus Fusarium oxysporum and involvement of cytochrome P-450 in the respiratory nitrite reduction. J Biol Chem. 1991; 266: 11078-11082.
- Morozkina EV, Kurakov AV. Dissimilatory nitrate reduction in fungi under conditions of hypoxia and anoxia: a review. Prikl Biokhim Mikrobiol. 2007; 43: 607-613.
- Bender D, Kaczmarek AT, Kuester S, Burlina AB, Schwarz G. Oxygen and nitrite reduction by heme-deficient sulphite oxidase in a patient with mild sulphite oxidase deficiency. J Inherit Metab Dis. 2020; 43: 748-757.
- Eh M, Kaczmarek AT, Schwarz G, Bender D. Molecular mechanism of intramolecular electron transfer in dimeric sulfite oxidase. J Biol Chem. 2022; 298: 101668.
- Dungel P, Teuschl AH, Banerjee A, Paier-Pourani J, Redl H, et al. Impact of mitochondria on nitrite metabolism in HL-1 cardiomyocytes. Front Physiol. 2013; 4: 101.
- Duvigneau JC, Kozlov AV. Pathological Impact of the Interaction of NO and CO with Mitochondria in Critical Care Diseases. Front Med (Lausanne). 2017; 4: 223.
- Donat ME, Wong K, Staines WA, Krantis A. Heme oxygenase immunoreactive neurons in the rat intestine and their relationship to nitrergic neurons. J Auton Nerv Syst. 1999; 77: 4-12.
- Hall ED, Wang JA, Bosken JM, Singh IN. Lipid peroxidation in brain or spinal cord mitochondria after injury. J Bioenerg Biomembr. 2016; 48: 169-174.
- Hill RL, Singh IN, Wang JA, Hall ED. Time courses of post-injury mitochondrial oxidative damage and respiratory dysfunction and neuronal cytoskeletal degradation in a rat model of focal traumatic brain injury. Neurochem Int. 2017; 111: 45-56.
- Kalbitz M, Waller C, Huber-Lang M, Gebhard F, Thiemermann C, et al. Oxygen in the Heart: How Much is too Much? Shock. 2017; 47: 531-532.
- Shaefi S, Talmor DS, Subramaniam B. Oxygen Therapy: When Is Too Much Too Much? Anesthesiology. 2016; 125: 449-450.
- Shaefi S, Marcantonio ER, Mueller A, Banner-Goodspeed V, Robson SC, et al. Intraoperative oxygen concentration and neurocognition after cardiac surgery: study protocol for a randomized controlled trial. Trials. 2017; 18: 600.
- Luc R, Vergely C. Forgotten radicals in biology. Int J Biomed Sci. 2008; 4: 255-259.
- Rutkowski R, Pancewicz SA, Rutkowski K, Rutkowska J. Reactive oxygen and nitrogen species in inflammatory process. Pol Merkur Lekarski. 2007; 23: 131-136.
- Sitdikova GF, Zefirov AL. Hydrogen sulfide: from the sewers of Paris to the signaling molecule. Priroda. 2010; 29-37.
- Sukmansky OI, Reutov VP. Gas transmitters: physiological role and participation in the pathogenesis of diseases. Uspekhi Fiziologicheskikh Nauk. 2016; 47: 30-58.
- Gusakova SV, Smagliy LV, Birulina Yu G, Kovalev IV, Nosarev V, et al. Molecular mechanisms of action of NO, CO and H2S gas transmitters in smooth muscle cells and the effect of NO-generating compounds (nitrates and nitrites) on average life expectancy. Uspekhi Fiziologicheskikh Nauk. 2017; 48: 24-52.
- Steiger C, Uchiyama K, Takagi T, Mizushima K, Higashimura Y, et al. Prevention of colitis by controlled oral drug delivery of carbon monoxide. J Control Release. 2016; 239: 128-136.
- Steiger C, Hermann C, Meinel L. Localized delivery of carbon monoxide. Eur J Pharm Biopharm. 2017; 118: 3-12.
- Ryter SW. Heme oxygenase-1/carbon monoxide as modulators of autophagy and inflammation. Arch Biochem Biophys. 2019; 678: 108186.
- Bhatia M. H2S and Inflammation: An Overview. Handb Exp Pharmacol. 2015; 230: 165-180.
- Yang R, Yu T, Liu D, Shi S, Zhou Y. Hydrogen sulfide promotes immunomodulation of gingiva-derived mesenchymal stem cells via the Fas/FasL coupling pathway. Stem Cell Res Ther. 2018; 9: 62.
- Reutov VP. Biochemical predetermination of the NO-synthase and nitrite reductase components of the nitric oxide cycle. Biochemistry. 1999; 64: 634-651.
- Reutov VP. Biomedical aspects of the cycles of nitric oxide and superoxide anion radical. Vestn RAMS. 2000; 35-41.
- Reutov VP. The nitric oxide cycle in mammals and the principle of cyclicity. Biochemistry. 2002; 67: 353-376.
- Reutov VP, Sorokina EG, Pinelis VG. Are nitrite ions involved in the regulation of intra- and intercellular signaling systems?. Vopr med chemistry. 1994; 40: 27-30.
- Lundberg JO, Gladwin MT, Ahluwalia A, Benjamin N, Bryan NS, et al. Nitrate and nitrite in biology, nutrition and therapeutics. Nat Chem Biol. 2009; 5: 865-869.
- Basu S, Azarova NA, Font MD, King SB, Hogg N, et al. Nitrite reductase activity of cytochrome c. J Biol Chem. 2008; 283: 32590-35297.
- Gladwin MT, Raat NJ, Shiva S, Dezfulian C, Hogg N, et al. Nitrite as a vascular endocrine nitric oxide reservoir that contributes to hypoxic signaling, cytoprotection, and vasodilation. Am J Physiol Heart Circ Physiol. 2006; 291: H2026-35.
- Appel KE, Schrenk D, Schwarz M, Mahr B, Kunz W. Denitrosation of N-nitrosomorpholine by liver microsomes; possible role of cytochrome P-450. Cancer Lett. 1980; 9: 13-20.
- Appel KE, Rühl CS, Hildebrandt AG. Metabolic inactivation of N-nitrosamines by cytochrome P-450 in vitro and in vivo. IARC Sci Publ. 1984; 57: 443-451.
- Amelizad S, Appel KE, Schoepke M, Rühl CS, Oesch F. Enhanced demethylation and denitrosation of N-nitrosodimethylamine by human liver microsomes from alcoholics. Cancer Lett. 1989; 46: 43-49.
- Turk JL. Rudolf Virchow – father of cellular pathology. J R Soc Med. 1993; 86: 688-689.
- Winau R. Rudolf Virchow and changes in medicine in the 19th Century. Verh Dtsch Ges Pathol. 1997; 81: 28-34.
- Leĭbson LG. L.A. Orbeli and various aspects of evolutionary physiology. Zh Evol Biokhim Fiziol. 1983; 19: 322-332.
- Kryzhanovsky G.N. Modern pathophysiology. Cas Lek Cesk. 1992; 131: 812-813.
- Kryzhanovskiĭ G.N. Pathophysiology as a modern integrative biomedical science. Vestn Akad Med Nauk SSSR. 1991; 3: 9-11.
- Reutov VP, Kayushin LP, Sorokina EG. Physiological role of the nitric oxide cycle in humans and animals. Human Physiology. 1994; 20: 165-174.
- Reutov VP, Azhipa Ya I, Kayushin LP. Oxygen as an inhibitor of hemoglobin nitrite reductase activity. Izv Acad of Sci of the USSR Ser boil. 1983; 3: 408-418.
- Cosby K, Partovi KS, Crawford JH, Patil RP, Reiter CD, et al. Nitrite reduction to nitric oxide by deoxyhemoglobin vasodilates the human circulation. Nat Med. 2003; 9: 1498-1505.
- Reutov VP, Shekhter AN How in the 20th century physicists, chemists and biologists answered the question: what is life? Physics– Uspekhi. 2010; 53: 377-396.
- Reutov VP, Sorokina EG, Pinelis VG Are nitrite ions involved in the regulation of intra- and intercellular signaling systems?. Vopr med chemistry. 1994; 40: 27-30.
- Reutov VP, Samosudova NV, Sorokina EG. Model of glutamate neurotoxicity and mechanisms of development of a typical pathological process. Biophysics. 2019; 64: 316-336.
- Hartley S, Dauvilliers Y, Quera-Salva MA. Circadian Rhythm Disturbances in the Blind. Curr Neurol Neurosci Rep. 2018; 18: 65.
- Liao G, Bose A. Entrainment within hierarchical circadian oscillator networks. Math Biosci. 2022; 351:108883.
- Yang W, Yuste R. Holographic imaging and photostimulation of neural activity. Curr Opin Neurobiol. 2018; 50: 211-221.
- Yang W, Yuste R. Holographic Imaging and Stimulation of Neural Circuits. Adv Exp Med Biol. 2021; 1293: 613-639.
- Szabo V, Ventalon C, De Sars V, Bradley J, Emiliani V. Spatially selective holographic photoactivation and functional fluorescence imaging in freely behaving mice with a fiberscope. Neuron. 2014; 84: 1157-1169.
- Pégard NC, Mardinly AR, Oldenburg IA, Sridharan S, Waller L, et al. Three-dimensional scanless holographic optogenetics with temporal focusing (3D-SHOT). Nat Commun. 2017; 8: 1228.
- Azhipa Ya I, Reutov VP, Kayushin LP. Ecological and medico-biological aspects of environmental pollution by nitrates and nitrites. Human Fiziology. 1993; 16: 220.
- Reutov VP, Davydova LA, Sorokina EG. Tissue-Engineered Constructions in Biophysics, Neurology and Other Fields and Branches of Medicine. Biophysics (Oxf). 2022; 67: 816-834.
